


ヒトだけ?「腸脛靭帯」の存在
ヒトの大腿部外側に位置する「腸脛靭帯(ITB: Iliotibial Band)」は、比較解剖学の視点から見ると極めて特異な構造です。チンパンジーをはじめとする現生の類人猿にはこの構造は存在せず、人類の系統においてのみ独自に獲得された「自己派生形質(autapomorphy)」であることが判明しています。
長年、ITBは二足歩行時に骨盤を側方から支える受動的なスタビライザーであると考えられてきました。しかし、近年のバイオメカニクス研究は、この伝統的な見解に疑問を投げかけています。本稿では、人類の移動様式の進化において、ITBが単なる支柱を超え、走行効率を劇的に高める「弾性エネルギー貯蔵庫」としていかに機能しているのか、その解剖学的真実を解き明かします。
人間の腸脛靭帯とチンパンジーの大腿筋膜
ヒトのITBは、類人猿が持つ大腿筋膜が特殊化したものです。最新の解剖学的データに基づき、その構造的な差異を比較します。

| 構造指標 | ヒトの腸脛靭帯 (ITB) | チンパンジーの大腿筋膜 (FL) |
| 主要な停止筋肉 | TFL, GMax (大臀筋) | TFL, GMaxCr (頭側), GMaxCd (尾側) |
| 筋線維の融合パターン | TFLとGMaxの大部分が合流し、強力な腱状の帯を形成 | TFLとGMaxCr前方部が遠位で融合。GMaxCdは主に大腿骨へ停止 |
| GMaxの停止様式 | 大臀筋の大部分がITBに停止 | GMaxCrの約5%、GMaxCdの約25%のみが筋膜へ停止 |
| 正規化PCSA (cm²/kg²/³) | TFL: 0.24 / GMax-ITB: 0.72 | TFL: 0.07 / GMaxCd-FL: 0.30 |
解剖学的な相同性に言及すると、人間の大臀筋はチンパンジーのGMaxCrと相同ですが、機能的なエネルギー貯蔵の観点からは、後方に位置するチンパンジーのGMaxCdと比較することが適切です。特筆すべきは、人間にはGMaxCd(尾側頭)が完全に欠損しており、代わりに巨大化した大臀筋の大部分がITBへと停止することで、腱膜を介した強力な力伝達を可能にしている点です。
生理学的断面積(PCSA)による分析
靭帯への荷重能力を決定するのは、停止する筋肉の「生理学的断面積(PCSA)」です。人間とチンパンジーの筋肉を正規化PCSA(体重を考慮した断面積)で比較すると、その力の生成能力には顕著な差があります。
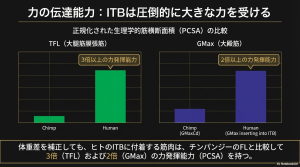
- 大腿筋膜張筋(TFL)
人間の正規化PCSAはチンパンジーの3倍以上であり、前方の靭帯成分を強力に牽引します。 - 大臀筋の伝達力
ITBに停止する人間側の大臀筋の正規化PCSAは、チンパンジーの対応部位(GMaxCd)の2倍以上に達します。
チンパンジーでは大臀筋の大部分が直接大腿骨に停止し、筋膜に伝達される力は限定的です。一方、人間は「筋肉の巨大化」と「ITBへの高い停止割合」という二重の進化により、靭帯を強力なスプリングとして活用する物理的強さを備えたのです。
腸脛靭帯のバイオメカニクス的意義
参考にした研究の定量的モデリングは、ITBが二足歩行において驚異的なエネルギー貯蔵能力を持つことを示しました。
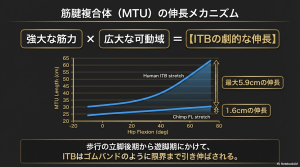
弾性能の定量的評価
ヒトのITBは、二足歩行時においてチンパンジーのFLよりも体重・歩幅あたり15〜20倍もの弾性エネルギーを蓄えることができます。さらに高速走行時において、ITBはアキレス腱が貯蔵するエネルギーの約14%に相当する量を蓄えており、これはランニングエコノミーに大きく寄与しています。
エネルギー貯蔵のメカニズム
- 前方ITB(TFL由来): 立脚後半から遊脚前半にかけて、大きなMTU(筋・腱複合体)が伸展によって引き伸ばされます。
- 後方ITB(GMax由来): ヒトとチンパンジーの比較において、後方のピーク歪み自体は同等ですが、人間は前述の通りPCSAが圧倒的に大きいため、伝達される荷重が格段に高く、結果として貯蔵されるエネルギー密度が劇的に向上します。
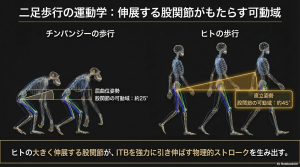
これは、類人猿の屈曲した歩行(crouched gait)から、人間特有の股関節を大きく伸展・屈曲させる直立二足歩行への移行が、ITBを「機能的なバネ」へと変貌させたことを意味します。
骨盤の安定化 vs 効率性の向上
ITBの主機能が「骨盤の側方安定化」であるとする従来の説は、近年のシミュレーションによって異議を唱えられています。

解析の結果、ITBが提供する外転モーメントは、股関節周囲の筋肉が持つ総外転モーメント能力の約10%に過ぎないことが判明しました。さらに、大臀筋の特定部位が骨盤を下方へ引く拮抗的なモーメントを生み出すことを考慮すると、その寄与はさらに限定的となります。
ITBは「安定」のためではなく、むしろ移動の効率性を高めるために選択された適応形質であると結論付けられます。
進化のタイムラインはアウストラロピテクスからホモ属へ
化石記録とバイオメカニクス的知見を統合すると、ITBの発達には明確なステップが存在したと推測されます。
- 直立姿勢の獲得(アウストラロピテクス)
約200万〜300万年前、坐骨の向きの変化や内反膝の獲得により、ITBを引き伸ばすための、幾何学的な配置になりました。しかし、この段階ではまだ筋肉量が不足しており、エネルギー貯蔵量は現代人には及びませんでした。 - 筋肉の巨大化(ホモ属)
ホモ・エレクトス以降、大臀筋の付着部が拡大し、ITBへ伝達される力が増大しました。これにより、現代的なITBの機能が完成しました。 - 持久狩猟への適応
最後に、これらの解剖学的変化が統合され、走行時のエネルギー消費を最小限に抑える「持続走行の要」としてのITBが確立されました。
最後共通祖先は、ナックルウォーク(ゴリラなどが手を使って歩く様式)を行うITBを持たない個体であった可能性が高く、ITBは人類が二足歩行に特化する過程で、新規に獲得された特殊構造なのです。
腸脛靭帯はヒトが長く移動するための武器
腸脛靭帯(ITB)は、単なる支持組織ではありません。それは、類人猿のエネルギー消費が激しい屈曲歩行から、人類の極めて効率的な直立移動へのシフトを象徴する解剖学的な特徴です。

大臀筋が生み出す強大な力を弾性エネルギーとして蓄え、再利用するこのシステムは、人類が広大な大地を移動し、狩猟で生き残るためのカギとなりました。私たちが今、長い距離を移動できるのは、ヒトの筋膜をスプリングへと変え、移動能力を極限まで高めた進化の結晶が、私たちの太ももに備わっているからなのです。


